
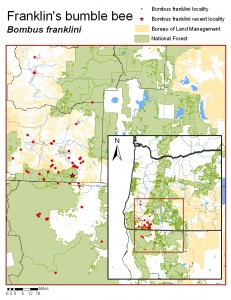
Bombus franklini occurs only in the USA. It is found only from southern Oregon to northern California between the Coast and Sierra-Cascade Ranges, in Douglas, Jackson, and Josephine in Oregon and Siskiyou and Trinity counties in California.
Bombus franklini is readily distinguished from other bumble bees in its range by the extended yellow on the anterior top of the thorax which extends well behind the wing bases and forms an inverted U-shape around the central patch of black, lack of yellow on the abdomen, predominantly black face with yellow on the vertex (top of the head), and white at the tip of the abdomen. Other bumble bees with similar color patterns in the range of B. franklini have the yellow extending back to the wing bases or only slightly beyond and usually have one or more bands of yellow on the middle or slightly behind the middle of the abdomen (most on tergum 4). Females of most species have yellow hair on the face, in contrast to black on B. franklini. Females of B. californicus which have black hair on the face also have black hair on the vertex in contrast to the yellow hair on the vertex in B. franklini. Additionally, B. californicus have yellow on the abdomen while B. franklini does not, and have a long face in contrast to the round face of B. franklini. Females of B. occidentalis are similar to B. franklini in the round faces, and black hair on the face, but differ in having the hair on the vertex black, as opposed to yellow. Additionally, females of B. occidentalis are variable in body hair color: some have yellow on the rear end of the thorax and the second and/or third abdominal segments, and even if the abdomen lacks yellow as in B. franklini, the pale hairs near the tip of the abdomen are more prominent in B. occidentalis.
Queens & workers of B. franklini are described as follows: face round with area between bottom of compound eye and base of mandible (= malar space) shorter than wide; hair predominantly black with some shorter light hairs intermixed above and below antennal bases. Hair on top of head (= vertex) yellow. Hair of thorax (= mesosoma) on anterior two-thirds above (= scutum) yellow extending rearward laterally inside and beyond the wing bases (= tegulae) to rear third (= scutellum), but interrupted medioposteriorly by inverted U-shaped patch of black; hair on posterior third (= scutellum) black above; hair of lateral thorax (= mesopleura) black, except for small patch of yellow in upper anterior corner in area of pronotal lobes. Hair of abdomen (= metasoma) black except for whitish or silvery hair at sides and apex of 5th plate above (= tergum 5, = T-5).
Males of this species are similar to females, differing as follows: malar space as long as wide, face below antennae with predominantly yellow hair, and tergum 6 with some pale hair laterally.
Technical keys, including color pattern illustrations of B. franklini and species that it might be confused with, are presented in Stephen (1957) and Thorp et al. (1983).
Bombus franklini is a primitively eusocial bumble bee. Like all other bumble bees, this species lives in colonies consisting of a queen and her immature and adult offspring: sterile female workers and males. The division of labor among the three types of adult bees is as follows. Queens are responsible for initiating colonies and laying eggs, workers are responsible for most food collection, colony defense, nest construction, and feeding of the young, and males sole function is to mate with new queens produced at the end of the colony season. Bumble bee colonies depend on floral resources for their nutritional needs; nectar provides carbohydrates for adult flight fuel and basic colony energy needs and pollen provides protein primarily for offspring growth.
The nesting biology of B. franklini is unknown, but, like other members of the subgenus Bombus sensu stricto, it probably nests underground in abandoned rodent burrows, or, occasionally, clumps of grass on the ground (Plath 1927; Thorp et al. 1983; Laverty and Harder 1988, Macfarlane et al. 1994, Hobbs 1968). Colonies are annual, initiated in spring by solitary, mated queens who emerge out of hibernation to search for appropriate nesting sites. The queen collects nectar and pollen from flowers to support the production of her eggs, which are fertilized by sperm she has stored since mating the previous fall. In the early stages of colony development, the queen is responsible for all food collection and care of the larvae. As the colony grows, the workers take over the duties of food collection, colony defense, nest construction, and larval care, while the queen remains within the nest and spends most of her time laying eggs. At their peak, colonies may consist of between 50 and 400 workers and the queen (Plath 1927; Thorp et al. 1983; Macfarlane et al. 1994). Two colonies of B. franklini initiated in the laboratory and set out to complete development in the field contained over 60 workers by early September, and probably produced over 100 workers by end of season (Plowright and Stephen 1980). Near the end of the colony cycle, reproductives (queens and males) are produced. Male bumble bees patrol selected territories, producing a queen-attracting scent which they deposit in suitable places and replace if it rains. When a female locates a male patrol-area, she remains still until a male finds her, and mating usually takes place on vegetation or the ground. Queens usually mate with only one male, but males may mate with multiple queens. After mating, the queens feed to build up their fat bodies, then enter hibernation for the winter. At the end of the colony season the founding queen, all workers and males die and the inseminated hibernating new queens are left to carry on the line into the following year.
In Bombus sensu stricto, queen production is complicated, apparently depending on a switch point when the first males are produced. This switch point usually occurs when queens cease producing a pheromone that inhibits workers from laying haploid (male producing) eggs, and is followed by a competition point when some workers initiate egg laying and compete with the queen for survival of their male progeny. An early switch point results in more males and fewer queens, while a late switch point results in larger colonies, more queens, and fewer males (Duchateau and Velthius 1988).
The flight season of B. franklini is from mid-May to the end of September (Thorp et al. 1983); a few individuals have also been encountered in October (Southern Oregon University Bee Collection records).
Franklin’s Bumble Bee has the most limited geographic distribution of any bumble bee in North America and possibly the world (Williams 1998). It is known only from southern Oregon and northern California between the Coast and Sierra-Cascade Ranges. Stephen (1957) recorded it from the Umpqua and Rogue River Valleys of Oregon. Thorp et al. (1983) recorded it from northern California and suggested its restriction to the Klamath Mountain region of southern Oregon and northern California. Its entire distribution, including historic populations and recent range extensions (Thorp 1999, 2001, 2004) can be covered by an oval of about 190 miles north to south and 70 miles east to west between 122o to 124o west longitude and 40o 58o to 43o 30o north latitude. It is known from Douglas, Jackson, and Josephine counties in Oregon and Siskiyou and Trinity counties in California. Elevations of localities where it has been found range from 540 feet (162 m) in the north to above 7800 feet (2340 m) in the south of its historic range. Although the number of populations that existed prior to 1998 is unknown, there are several historical records for this species, both published and in museums, including two in 1925 (Gold Hill and Roseburg, OR), one in 1930 (Roseburg, OR), two in 1950 (Gold Hill and Medford, OR), two in 1958 (Ashland, OR), two in 1968 (Mt. Ashland and near Copper, OR), one in 1980 (Ashland, OR), two in 1988 (Ashland and Merlin, OR), two in 1989 (Hilt and Yreka, CA), four in 1990 (Ashland, Ruch, Central Point, and Gold Hill, OR), one in 1992 (Ashland, OR), two in 1997 (Roxy Ann Peak near Medford and Ashland Pond in Ashland, OR), and four in 1998 (Roca Canyon in Ashland, Lost Creek Reservoir, and Grizzly Peak near Shale City, OR). Additional records with unknown dates and or localities are also available, including the 1917 type specimen whose locality (Nogales, AZ) has been determined to be erroneous. Post-1998, the numbers of populations and individuals of B. franklini have declined drastically, evidenced by intensive and extensive surveys primarily conducted by R. W. Thorp each year from 1998 through 2008 (Thorp 1999, 2001, 2004, 2005b, c, 2008). During each of the past 11 years, Dr. Thorp surveyed from nine to 17 historic sites (average 12.3 sites) some visited more than once each year with an average of 29.6 visits to historic sites per year. Dr. Thorp also made from two to 23 visits to additional sites with potential habitat for B. franklini each year (averaging 12.0 site visits per year), with some sites visited more than once per year and some visited in multiple years. Additional surveys in 2006 were conducted by the BLM; 16 sites with optimal habitat for B. franklini were each surveyed twice (Code & Haney 2006).
Between 1998 and 2005, the number of sightings of Bombus franklini declined precipitously from 94 individuals in 1998 to 20 in 1999, nine in 2000 and one in 2001. Although 20 were found in 2002 only three were sighted in 2003, all at a single locality. None were found in 2004 and 2005, and a single worker was sighted in 2006. This was at the same locality, Mt. Ashland, where the last B. franklini were found in 2003. No individuals were found in 2007 or 2008.
No B. franklini were found during the BLM search effort in 2006 (Code & Haney 2006).
B. franklini requires habitat with a sufficient supply of floral resources to provide continuous blooming throughout the colony season. Bumble bees are generalist foragers, gathering pollen and nectar from a wide variety of flowering plants. Bombus franklini have been observed collecting pollen on lupine (Lupinus) and California poppy (Eschscholzia), and nectaring on horsemint (Agastache) and mountain penny-royal (Monardella) (R. Thorp pers. obs.). They may collect both pollen and nectar from vetch (Vicia) and also rob nectar from this plant (P. Schroeder 2006 pers. comm.).
Landscape level habitat quality has been shown to influence bumble bee species richness and abundance, indicating that isolated patches of habitat are not sufficient to fully support bumble bee populations (Hatfield & LeBuhn 2007; Öckinger & Smith 2007). Additionally, since B. franklini probably requires abandoned rodent borrows or clumps of grass for nesting, population sites may be limited by the abundance of rodents and the presence of undisturbed grassland.
Bombus franklini is currently considered to be a Species of Concern by the U.S. Fish and Wildlife Service (USFWS 2009). Species of Concern are defined as taxa whose conservation status is of concern to the U.S. Fish and Wildlife Service (many previously known as Category 2 candidates), but for which further information is still needed. Such species receive no legal protection and use of the term does not necessarily imply that a species will eventually be proposed for listing (ODFW 2009). Bombus franklini is also included on the California Department of Fish and Game special animals list (CDFA 2009) and is listed as an interagency special status species by the USDA Forest Service and Bureau of Land Management (Forest Service/BLM 2009). The species has a NatureServe Global Status rank of G1, an Oregon state status rank of S1 and a California state status rank of SNR (NatureServe 2009). It is listed as critically imperiled by the IUCN Red List of endangered species (IUCN 2009) and the Red List of Pollinator Insects of North America, produced by the Xerces Society for Invertebrate Conservation (Thorp 2005a).
Bombus franklini (Franklin’s Bumble Bee) is in imminent danger of extinction. Extensive surveys from 1998-2008 have demonstrated that there has been a precipitous decline in the number of individuals and localities in the past decade. In 1998, 94 individuals were found at 8 sites, while in the past three years, only one individual has been observed in surveys. The threats to this species are outlined as follows:
Disease: The recent declines of Bombus franklini and its close relatives are hypothesized to be primarily due to a selectively virulent strain of the microsporidian Nosema bombi acquired from the commercially reared and closely related European Large Earth Bumble Bee, Bombus terrestris (Thorp 2003, Thorp & Shepherd 2005, National Research Council 2007). Commercially reared bumble bees frequently harbor pathogens and their escape from greenhouses can lead to infections in native species (Colla et al. 2006, Ottersttatter and Thomson 2008, Durrer & Schmid-Hempel 1994, Goka et al. 2001, 2002). Other pests and diseases found in commercial bumble bees that could be leading to the continued decline in North American bumble bees include the protozoan parasite Crithidia bombi, the tracheal mite Locustacarus buchneri, and the RNA virus known as Deformed Wing Virus (DWV).
Habitat Alteration: Any habitat alteration which destroys, fragments, degrades, or reduces food supplies, nest sites, or over-wintering sites can harm this species. In particular, agricultural intensification, livestock grazing, urban development, and road construction can fragment bumble bee habitat into pieces that are too small or too distant to support diverse bumble bee communities (Goulson et al. 2008). Fragmented habitats may not support healthy metapopulation structures and may eliminate or decrease source populations of bumble bees for recolonization (National Research Council 2007). Additionally, as small, isolated populations get smaller and more separated from adjacent populations, the pool of local genetic material shrinks and breeding between closely related individuals can result in inbreeding depression, a fitness reduction which lowers the population’s ability to survive and reproduce. Inbred populations of bumble bees show decreased genetic diversity and are at a greater risk of decline (Zayed and Packer 2005; Darvill et al. 2006; Ellis et al. 2006). Small or inbred populations may also lose resilience to environmental changes, and are at risk of extirpation due to stochastic events such as fire or unusually wet or dry years.
Pesticide Applications: The National Academy of Science National Research Council’s report on the Status of Pollinators in North America notes that bumble bees can be negatively affected by many pesticides and that ground-nesting bumble bees are uniquely susceptible to pesticides that are used on lawns or turf (National Research Council 2007). Foraging bees are poisoned by pesticides when they absorb the fast-acting toxins through their integument (the outer “skin” that forms their exoskeleton), drink contaminated nectar, or gather pesticide-covered pollen or micro-encapsulated poisons. Pesticide drift from aerial spraying can kill 80% of foraging bees close to the source and drift can continue to be dangerous for well over a mile (Johansen and Mayer 1990). Insecticide application on Forest Service managed public lands for spruce budworm has been shown to cause massive kills of bumble bees and reduce pollination of nearby commercial blueberries in New Brunswick (reviewed in Kevan & Plowright 1995). Insecticides applied in the spring when bumble bee queens are foraging and colonies are small are likely to have a more significant effect on bumble bee populations than insecticides that are applied at other times of the year (Goulson et al. 2008). The relatively recent and increased use of persistent neonicotinoid pesticides, known to be highly toxic to bees, may pose an increased threat to bumble bees in the subgenus Bombus (Colla and Packer 2008). These and other pesticides may be translocated through plants into the nectar or pollen. In addition to insecticides, broad-spectrum herbicides used to control weeds can indirectly harm bumble bees by removing the flowers that would otherwise provide the bees with pollen and nectar (Williams 1986; Shepherd et al. 2003, Smallidge & Leopold 1997). Bumble bees require consistent sources of nectar, pollen, and nesting material during the adult activity period, and reduction of these resources by herbicides can cause a decline in bumble bee reproductive success and/or survival rates. For example, Kevan (1999) found that herbicide applications have reduced the reproductive success of blueberry pollinators by limiting alternative food sources that can sustain the insects when the blueberries are not in bloom.
Fire suppression and natural/introduced fire: Although fire is an important natural and managed disturbance throughout natural areas in the United States, suppression of wildfire is seen as necessary to protect natural resources, homes, and businesses. Fire suppression can lead to extensive changes in vegetation structure, including degradation and loss of grasslands and herbaceous species as the shrub community matures (Panzer 2002, Schultz & Crone 1998). Continued fire suppression not only results in the reduction of bumble bee food plants at these sites, but also renders the habitat susceptible to catastrophic, large scale, and high temperature fires due to increases in combustible fuel loads, tree density, and fire intolerant species (Huntzinger 2003). Controlled burns, however, could inadvertently threaten this species, and if executed, must be done so only with extreme caution, proper timing, and expert advice.
Invasive and introduced species: Invasion and dominance of native grasslands by exotic plants is a common issue (Warren 1993, Schultz 1998), and has likely occurred at historic B. franklini locations. Introduced plants could threaten B. franklini by directly competing with plants providing nectar and pollen.
Global climate change: A changing climate may cause shifts in the range of host plant species available to pollinators, and can be especially detrimental to pollinators when combined with habitat loss (National Research Council 2007). Although the very large size of the queens suggests that the dispersal ability of this species may be greater than that of other bumble bees within its range, the patchy distribution of its remaining habitat might hinder dispersal made necessary by climate change. Additionally, increased amounts of UV-B radiation from a reduction in ozone could delay flowering in plants and reduce the amount of flowers that plants produce (National Research Council 2007), while an increase in atmospheric CO2 from global climate change may alter plant nectar production, all of which could have negative effects on the foraging success of bumble bees (reviewed in Davis 2003).
Competition from honey bees (Apis mellifera): It has long been assumed, but difficult to demonstrate, that managed honey bee colonies have a negative impact on native bees through competition for floral resources (Sugden et al. 1996; Butz Huryn 1997). Recently, Thomson (2004, 2006) conducted competition experiments on B. occidentalis colonies placed at three distances from introduced honey bee hives. Thomson found decreased foraging activity, especially for pollen, and lowered reproductive success in Bombus occidentalis colonies nearest the Apis hives. Evans (2001) found the same results in a similar study with B. impatiens colonies in Minnesota. Goulson and Sparrow (2009) found that bumble bees are smaller, and thus less fit, in areas where managed honey bees occur in Scotland.
Although B. franklini is widely recognized as a vulnerable species, it receives no formal or informal protection. Since many plants rely on bumble bees to achieve pollination, their population declines and losses have far ranging ecological impacts. An examination of the theoretical effect of removal of specialist and generalist pollinators on the extinction of plant species concluded that the loss of generalist pollinators poses the greatest threat to pollinator networks (Memmott et al. 2004). In Britain and the Netherlands, where multiple bumble bee species, as well as other bees, have gone extinct, there is evidence of decline in the abundance of insect pollinated plants (Biesmeijer et al. 2006).
The primary threat to this species is the spread of disease by bumble bees from outside of its geographic range. There are currently no federal regulations that limit the interstate transport of bumble bees even outside their native range (Flanders et al. 2003).
Management: All known Bombus franklini habitat should be protected and managed to include plentiful food (pollen and nectar resources such as Lupinus, Eschscholzia, Agastache, Monardella, Vicia), abandoned rodent burrows in which to nest, and proximity to water sources (lakes, rivers, streams, seeps) for prolongation of flowering season of plant food sources. All efforts should be made to prevent the spread of disease from commercially reared and managed bumble bee colonies to native populations.
Research: Monitor, sample, and conduct population estimates of extant populations of B. franklini. Conduct research aimed at increasing understanding of the pathology and control of Nosema bombi [Microsporidia] and other potential disease organisms (such as Locustacrus buchneri [Acarina] and Crithidia bombi [Protozoa]). Focus research on elucidating the virulence and cross-infectivity of strains of these disease organisms, especially Nosema bombi, between commercially reared and wild bumble bee species in order to better assess the ecological risks of trafficking managed crop pollinators.
Biesmeijer, J. C., S. P. M Roberts, M. Reemer, R. Ohlemiller, M. Edwards, T. Peeters, A. P. Schaffers, S. G. Potts, R. Kleukers, C. D. Thomas, J. Settele, and W. E. Kunin. 2006. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 313(5785): 351-354.
Butz Huryn, V. 1997. Ecological impacts of introduced honey bees. Quarterly Review of Biology. 72: 275-297.
CDFA. 2009. California Department of Food and Agriculture, State of California, The Resources Agency, Department of Fish and Game. Biogeographic Data Branch: California Natural Diversity Database: Special Animals (901 taxa) February 2009. http://www.dfg.ca.gov/biogeodata/cnddb/pdfs/spanimals.pdf (Last accessed April 2009)
Code, B. H. and L. H. Stephan. 2006. Final Report: Franklin’s Bumble Bee Inventory in the Southern Cascades of Oregon. Prepared for: The Bureau of Land Management, Medford, Oregon, 4 Oct. 2006.
Colla, S. R. and L. Packer. 2008. Evidence for the decline of Eastern North American Bumble Bees, with special focus on Bombus affinis Cresson. Biodiversity and Conservation 17: 1379-1391.
Colla, S. R., M. C. Otterstatter, R. J. Gegear, and J. D. Thomson. 2006. Plight of the bumble bee: Pathogen spillover from commercial to wild populations. Biological Conservation 129: 461-467.
Darvill, B., J. S. Ellis, G. C. Lye, and D. Goulson. 2006. Population structure and inbreeding in a rare and declining bumblebee, Bombus muscorum (Hymenoptera: Apidae). Molecular Ecology 15: 601-611.
Davis, A. R. 2003. Influence of elevated CO2 and ultraviolet-B radiation levels on floral nectar production: a nectary-morphological perspective. Plant Systematics and Evolution 238: 169-181.
Duchateau M. J. and H. H. W. Velthuis. 1988. Development and reproductive strategies in the bumble bee, Bombus terrestris. Behaviour 107: 186-207.
Durrer, S. and P. Schmid-Hempel. 1994. Shared use of flowers leads to horizontal pathogen transmission. Proceedings of the Royal Society: Biological Sciences 258: 299-302.
Ellis, J. S., M. E. Knight, B. Darvill, and D. Goulson. 2006. Extremely low effective population sizes, genetic structuring and reduced genetic diversity in a threatened bumblebee species, Bombus sylvarum (Hymenoptera: Apidae). Molecular Ecology 15: 4375-4386.
Evans, E. C. 2001. Competition between European honey bees and native bumblebees: resource overlap and impact on reproductive success. Master’s Thesis, University of Minnesota.
Flanders, R. V., W. F. Wehling, and A. L. Craghead. 2003. Laws and regulations on the import, movement and release of bees in the United States. pp. 99-111. In K. Strickler and J. H. Cane (eds.) For Nonnative Crops, Whence Pollinators of the Future? Thomas Say Publications in Entomology: Proceedings. Entomological Society of America, Lanham, MD.
Forest Service/BLM. 2009. Interagency Special Status Sensitive Species Program. http://www.fs.fed.us/r6/sfpnw/issssp/species-index/fauna-invertebrates.shtml (Last accessed 27 April 2009).
Frankie, G. W., R. W. Thorp, M. Schindler, J. Hernandez, B. Ertter, and M. Rizzardi. 2005. Ecological patterns of bees and their host ornamental flowers in two northern California cities. Journal of the Kansas Entomological Society 78: 227-246.
Goka, K., K. Okabe, S. Niwa, and M. Yoneda. 2000. Parasitic mite infection in introduced colonies of European bumblebees, Bombus terrestris. Japanese Journal of Applied Entomology and Zoology 44: 47-50 (in Japanese with English abstract).
Goka, K., K. Okabe, M. Yoneda, and S. Niwa. 2001. Bumblebee commercialization has caused international migration of parasitic mites. Molecular Ecology 10: 2095-2099.
Goulson, D. 2003. Bumblebees, Their Behaviour and Ecology. Oxford University Press, Oxford.
Hatfield, R. G. and G. LeBuhn. 2007. Patch and landscape factors shape community assemblage of bumblebees, Bombus spp. (Hymenoptera: Apidae), in montane meadows. Biological Conservation 139: 150-158.
Hobbs, G.A. 1968. Ecology of species of Bombus Latr. (Hymenoptera: Apidae) in southern Alberta. VI. Subgenus Bombus. Canadian Entomologist 100: 156-164.
Huntzinger, M. 2003. Effects of fire management practices on butterfly diversity in the forested western United States. Biological Conservation 113: 1-12.
IUCN. 2009. Red List of Threatened Species. http://www.iucnredlist.org/details/135295. (Last accessed 26 Feb. 2009).
Johansen, C. A. and D. F. Mayer. 1990. Pollinator Protection: a Bee and Pesticide Handbook. Wicwas Press, Cheshire, Connecticut.
Kevan, P. G. 1999. Pollinators as bioindicators of the state of the environment: species, activity and diversity. Agriculture Ecosystems and Environment 74: 373-393.
Kevan, P. G. and R. C. Plowright. 1995. Impact of pesticides on forest pollination. pp. 607-618 In J. A. Armstrong and W. G. H. Ives (eds.) Forest Insect Pests in Canada. Natural Resources Canada, Canadian Forest Service, Science and Sustainable Development Directorate, Ottawa, Ontario, Canada.
Laverty, T. and L. D. Harder. 1988. The bumble bees of eastern Canada. Canadian
Entomologist 120: 965-987.
Macfarlane, R. P., K. D. Patten, L. A. Royce, B. K. W. Wyatt, and D. F. Mayer. 1994. Management potential of sixteen North American bumble bee species. Melanderia 50: 1-12.
Memmott, J., N. M. Waser, and M. V. Price. 2004. Tolerance of pollination networks to species extinctions. Proceedings of the Royal Society B 271: 2605-2611.
National Research Council. 2007. Status of Pollinators in North America. The National Academies Press, Washington, DC.
NatureServe. 2009. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. Available http://www.natureserve.org/explorer. (Last accessed 26 Feb. 2009).
Oregon Department of Fish and Wildlife (ODFW). 2009. http://www.fws.gov/oregonfwo/Species/Data/ (Last accessed 6 April 2009).
Öckinger, E. and H. G. Smith. 2007. Semi-natural grasslands as population sources for pollinating insects in agricultural landscapes. Journal of Applied Ecology 44: 50-59.
Otterstatter, M. C., and J. D. Thomson. 2008 Does pathogen spillover from commercially reared bumble bees threaten wild pollinators? PLoS ONE 3: e2771.doi:10.1371/journal.pone.0002771
http://www.plosone.org/doi/pone.0002771
Panzer, R. 2002. Compatibility of prescribed burning with the conservation of insects in small isolated prairie reserves. Conservation Biology 16 (5): 1296-1307.
Plath, O. E. 1927. Notes on the nesting habits of some of the less common New England bumblebees. Psyche 34: 122-128.
Schroeder, P. 2006. Personal communication with Robbin Thorp.
Schultz, C. B. 1998. Ecology and Conservation of the Fender’s Blue Butterfly. PhD. Dissertation, University of Washington, Seattle, WA.
Schultz, C. B. and E. E. Crone. 1998. Fire to restore butterfly habitat? A modeling approach to management tradeoffs for the Fender’s blue. Restoration Ecology 6: 244-252.
Shepherd, M., S. L. Buchmann, M. Vaughan, and S. H. Black. 2003. Pollinator Conservation Handbook. The Xerces Society, Portland, Oregon.
Smallidge, P. J. and D. J. Leopold. 1997. Vegetation management for the maintenance and conservation of butterfly habitats in temperate human-dominated habitats. Landscape and Urban Planning 38: 259-280.
Stephen, W. P. 1957. Bumble Bees of Western America. Oregon State College, Agricultural Experiment Station, Technical Bulletin 40, 163pp.
Sugden, E. A. 1985. Pollinators of Astragalus monoensis Barneby (Fabaceae) – New host records – Potential impact of sheep grazing. Great Basin Naturalist 45: 299-312.
Thomson, D. 2004. Competitive interactions between the invasive European honey bee and native bumble bees. Ecology 85: 458-470.
Thomson, D. M. 2006. Detecting the effects of introduced species: a case study of competition between Apis and Bombus. Oikos 114: 407-418.
Thorp, R. W., D. S Horning and L. L. Dunning. 1983. Bumble bees and cuckoo bumble bees of California (Hymenoptera: Apidae). Bulletin of the California Insect Survey 23: viii.
Thorp, R. W. 1999. Franklin’s bumble bee, Bombus franklini (Frison 1921): a species of special concern. Report to USDA Forest Service, Ashland. OR on 1998 season (submitted 23 Nov 99)
Thorp, R. W. 2001. Franklin’s bumble bee, Bombus franklini (Frison 1921): a species of special concern. Report to US Fish and Wildlife Service, Portland, OR on 1999-2000 seasons (submitted 14 May 2001)
Thorp, R. W. 2003. Bumble bees (Hymenoptea: Apidae): commercial use and environmental concerns. Pp. 21-40. In: K. Strickler and J. H. Cane (eds.). For nonnative crops, whence pollinators of the future? Thomas Say Publications in Entomology: Proceedings. Entomological Society of Amererica, Lanham, MD., 204pp.
Thorp, R. W. 2004. Franklin’s bumble bee, Bombus (Bombus) franklini (Frison). Report to US Fish and Wildlife Service, Portland, OR on 2001-2003 seasons (submitted 29 June 2004).
Thorp, R. W. 2005a. Species Profile: Bombus franklini. In Shepherd, M. D., D. M. Vaughan, and S. H. Black (Eds). Red List of Pollinator Insects of North America. CD-ROM Version 1 (May 2005). Portland, OR: The Xerces Society for Invertebrate Conservation.
Thorp, R. W. 2005c. Franklin’s bumble bee, Bombus (Bombus) franklini (Frison). Report to US Fish and Wildlife Service, Portland, OR on 2004 season (submitted 29 March 2005).
Thorp, R. W. 2005b. Franklin’s bumble bee, Bombus (Bombus) franklini (Frison). Report to US Fish and Wildlife Service, Portland, OR on 2005 season (submitted 7 November 2005).
Thorp, R. W. and Shepherd, M. D. 2005. Subgenus Bombus. Latreille, 1802 (Apidae: Apinae: Bombini). In Shepherd, M. D., D. M. Vaughan, and S. H. Black (eds.) Red List of Pollinator Insects of North America. CD-ROM Version 1 (May 2005). Portland, OR: The Xerces Society for Invertebrate Conservation. www.xerces.org/Pollinator_Red_List/Bees/Bombus_Bombus.pdf
USFWS, Oregon Fish and Wildlife Office. 2009, Endangered Species “Species Data” http://www.fws.gov/oregonfwo/Species/Data/. (Last accessed 26 Feb. 2009).
Warren, M. S. 1993. A review of butterfly conservation in central southern Britain: II. site management and habitat selection of key species. Biological Conservation 64: 37-49.
Williams, P. H. 1986. Environmental change and the distribution of British bumble bees (Bombus Latr.). Bee World 67: 50-61.
Williams, P. H. 1998. An annotated checklist of bumble bees with an analysis of patterns of description (Hymenoptera: Apidae, Bombini). Bulletin of the Natural History Museum, London (Ent.) 67: 79-152. updated at: www.nhm.ac.uk/research-curation/projects/bombus/
Zayed, A. and L. Packer. 2005. Complementary sex determination substantially increases extinction proneness of haplodiploid populations. Proceedings of the National Academy of Sciences of the United States of America 102 (30): 10742-10746.
Original profile prepared by Dr. Robbin Thorp, University of California- Davis. This profile has been updated by Sarah Foltz, Scott Black, Sarina Jepsen, and Elaine Evans, The Xerces Society.




